







With this fiber-electrode manipulator a broad range of even very thin shafted probes can be handled, including fiber and wire-electrodes with shaft diameters down to about 25 µm. The patented driving principle of the Eckhorn Manipulator offers an outstanding electrode positioning accuracy. Compared with hydraulic, direct motor driven or cable controlled microdrives, the patented fiber electrode manipulator “System Eckhorn” does not cause hysteresis errors of the electrode movement. Hysteresis error is generally a result of stiction and free motion of the driving mechanism. Our system has a higher degree of positioning accuracy due to its rubber tube driving mechanism being almost absolutely free of stiction and free motion.
 Figure 1: 7 Electrode (left) and 16 Electrode Eckhorn Matrix (right)
Figure 1: 7 Electrode (left) and 16 Electrode Eckhorn Matrix (right)
 Figure 2: Components of a 7 Electrode Eckhorn Matrix (without xyz-manipulator). In this case the Eckhorn Matrix is equipped with a rod to mount the drive to a stereotaxic instrument.
Figure 2: Components of a 7 Electrode Eckhorn Matrix (without xyz-manipulator). In this case the Eckhorn Matrix is equipped with a rod to mount the drive to a stereotaxic instrument.
 Figure 3: 7 Electrode Eckhorn Matrix mounted to a XYZ-manipulator
Figure 3: 7 Electrode Eckhorn Matrix mounted to a XYZ-manipulator
Using Thomas Eckhorn Matrix systems with patented rubber tube drive allows to record neural activity while the electrode is moving in the brain. So it is very easy to search active neurons in the brain.
The Thomas Eckhorn Matrix systems have some advantages in comparison to other multielectrode microdrive systems for neurophysiological research presently available on the market:
- Vibration-free positioning of up to 16 quartz-platinum/tungsten microelectrodes in 1µm steps
- Patented rubber tube driving principle
- High positioning accuracy, due to lack of stiction and free motion
- Easy exchange and calibration of microelectrodes within minutes
- Advances up to 16 microelectrodes/tetrodes independently in 1µm steps up to 35.000µm (other electrode travel distances on request!)
- Xyz-manipulator travel-distance: z=0-30mm, x= ±10mm, y=±10mm
- Continuous low noise recording while the electrode scans the tissue at low velocity,
- No noise introduction in the recordings so that you can search active neurons in the brain
- No electrode connection cables free in air, no electrical noise pickup from environment (e.g. motors, radio stations)! Competitor drives pickup electrical interference from the environment so that you for example can hear radio station music or electrical artifacts from lab devices from the audio monitor loudspeaker instead of neural spikes!
- Electrode moving velocity adjustable (1-250µm/s), moving direction selectable
- Low noise 7 or 16 channel preamplifier is integrated into the microdrive chassis, system is completely shielded to avoid electrical noise pickup from the environment, three preamplifier operation modes: record, electrode impedance test, electrical stimulation/lesioning
- System is completely delivered with motor control device, integrated preamplifier, software, xyz-manipulator, chamber holder, exchangeable head, handheld remote control, etc.
- Exchangeable manipulator head allows manifold electrode configurations and interelectrode spacings
- Electrode spacings from 80µm up to some millimeters possible due to exchangeable Eckhorn Matrix heads
- The system is modular and adaptable to the end user´s requirements.
- The Eckhorn Matrix is usable for cortical as well as deep brain recordings in all kinds of research applications
- User friendly computer controlled system with LAN (local area network) communication via TCP/IP protocol between motor control unit and personal computer. Software package is part of the system.
- Using very thin shaft Thomas microelectrodes (outer diameter 80µm) causes minimal tissue damage.
- Different neurophysiological methods are available for the Eckhorn Matrix systems (e.g. optogenetics, microinjection)
 Figure 4: Neural data recorded in a primate amygdala by using a 7 electrode Eckhorn Matrix (data kindly provided by Prof. Dr. Katalin Gothard, Arizona Health Science Center, Tucson, USA)
Figure 4: Neural data recorded in a primate amygdala by using a 7 electrode Eckhorn Matrix (data kindly provided by Prof. Dr. Katalin Gothard, Arizona Health Science Center, Tucson, USA)
The Eckhorn Matrix system is available for small animal stereotaxic instruments (see figure 10) as well as for primate experiments (see figure 11).
 Figure 5: Degrees of freedom of the 7 Electrode (left) and 16 Electrode Eckhorn Matrix XYZ manipulators
Figure 5: Degrees of freedom of the 7 Electrode (left) and 16 Electrode Eckhorn Matrix XYZ manipulators
Exchangeable Heads
 Figure 6: 16 Electrode Eckhorn Matrix with Xyz-Manipulator and exchangeable microdrive head
Figure 6: 16 Electrode Eckhorn Matrix with Xyz-Manipulator and exchangeable microdrive head
Thomas RECORDING offers a broad range of exchangeable Microdrive heads with different guide tube arrangements and electrode spacings for the 7- and 16-Electrode Eckhorn Systems.
 Figure 7: 16 Channel head with 4 individual adjustable groups of electrodes 4x(2×2)
Figure 7: 16 Channel head with 4 individual adjustable groups of electrodes 4x(2×2) Figure 8: 16 Channel head with 4×4 offset electrode arrangement
Figure 8: 16 Channel head with 4×4 offset electrode arrangement
There is a broad range of different Microdrive heads available. We also can offer customized solutions!
Electrical Stimulation / Impedance Test with the Eckhorn Matrix
The Eckhorn Matrix preamplifier is integrated in the microdrive chassis and has three operation modes: Recording / Electrode Impedance Test /Stimulation or Lesioning. The standard operation mode of the preamplifier is “Recording”. To use the other two modes Thomas RECORDING offers a special designed device that is called Mode Selection & Impedance Test Device (MSD).
The MSD is optional available, for more information please see the productpage of MSD.
 Figure 9: Mode Selection & Impedance Test Device for 7- or 16-channel Eckhorn Systems (1)=MSD, (2)=Control Box
Figure 9: Mode Selection & Impedance Test Device for 7- or 16-channel Eckhorn Systems (1)=MSD, (2)=Control Box
Thomas RECORDING offers a broad range of solutions for mounting the Eckhorn multielectrode systems to stereotaxic instruments. Below you can find some examples for Eckhorn microdrives mounted to small (see fig. 10) and large animal stereotaxic instruments (see figure 11). It is our strength to adapt our Microdrive to stereotaxic setups. Please feel free to ask for your individual adaptation.
 Figure 10: 32 Electrode Eckhorn Matrix mounted to a Thomas RECORDING small animal stereotaxic instrument with a special halo holder.
Figure 10: 32 Electrode Eckhorn Matrix mounted to a Thomas RECORDING small animal stereotaxic instrument with a special halo holder.
 Figure 11: 16 Electrode Eckhorn Matrix mounted to a Thomas RECORDING primate stereotaxic instrument (PPS=Precision Positioning System) with a special halo holder.
Figure 11: 16 Electrode Eckhorn Matrix mounted to a Thomas RECORDING primate stereotaxic instrument (PPS=Precision Positioning System) with a special halo holder.
For making drug injection experiments with the Thomas Eckhorn Matrix system we offer a special designed high pressure microinjection system. One of the 7- or 16-single microelectrodes of the Eckhorn Matrix can be replaced by a microinjection cannula. The microinjection cannula (OD=120µm) has a tapered tip and is equipped with the patented Thomas rubber tube drive so that you can position the microinjection cannula tip together with the recording microelectrodes with high positioning accuracy (better than 1µm). The microinjection system consists of a special designed and computer controlled syringe pump that is connected to the microinjection cannula via a special thick wall tubing.
 Figure 12: Microinjection pump (A), connection of thick wall tubing to syringe cannula (B)
Figure 12: Microinjection pump (A), connection of thick wall tubing to syringe cannula (B)
The precision micro motor of the microinjection pump is controlled by the same motor control unit and graphical user interface as the Thomas Eckhorn Matrix. This guarantees a high precision and convenient experimental control of the injection/recording experiment.
Thomas RECORDING offers a complete range of Eckhorn Matrix accessories for optogenetic experiments. One of the 7- or 16 single microelectrodes or tetrodes is replaced by an optical fiber (OD=120µm). The optical fiber has a tapered tip and is equipped with the patented Thomas rubber tube drive so that you can position the optical fiber tip together with the recording microelectrodes with high positioning accuracy (better than 1µm). Figure 13 shows an Eckhorn Matrix head with concentric guide tube arrangement and a single optical fiber in the center position. The recording microelectrodes are concentrically arranged around the optical fiber in the center of this arrangement.
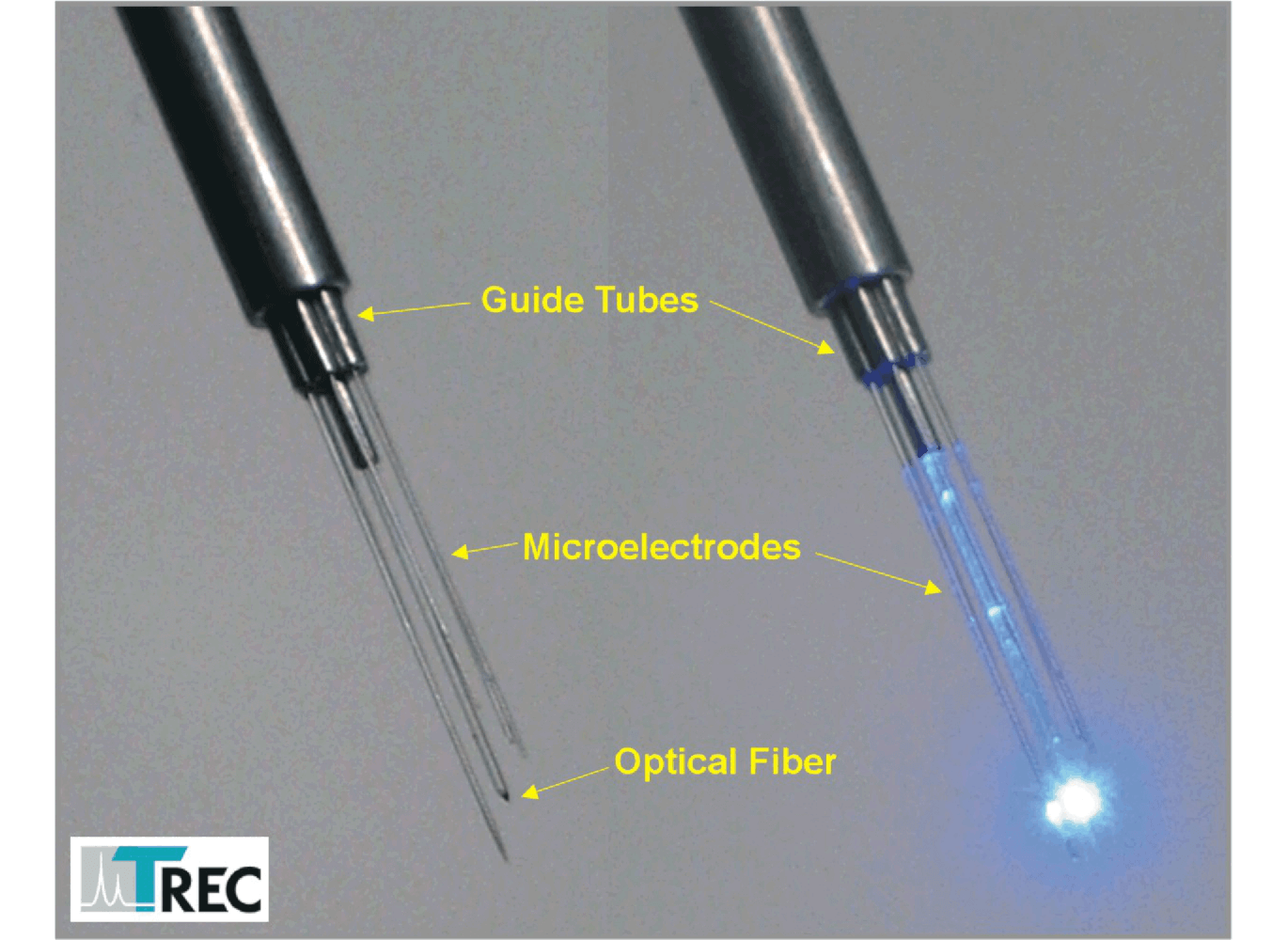 Figure 13: 7 Electrode Eckhorn Matrix concentric head with one optical fiber and 6 recording electrodes. (see [24])
Figure 13: 7 Electrode Eckhorn Matrix concentric head with one optical fiber and 6 recording electrodes. (see [24])

NEWS

PRODUCTS

SOLUTIONS

DISTRIBUTORS
 Sign Up
Sign Up