



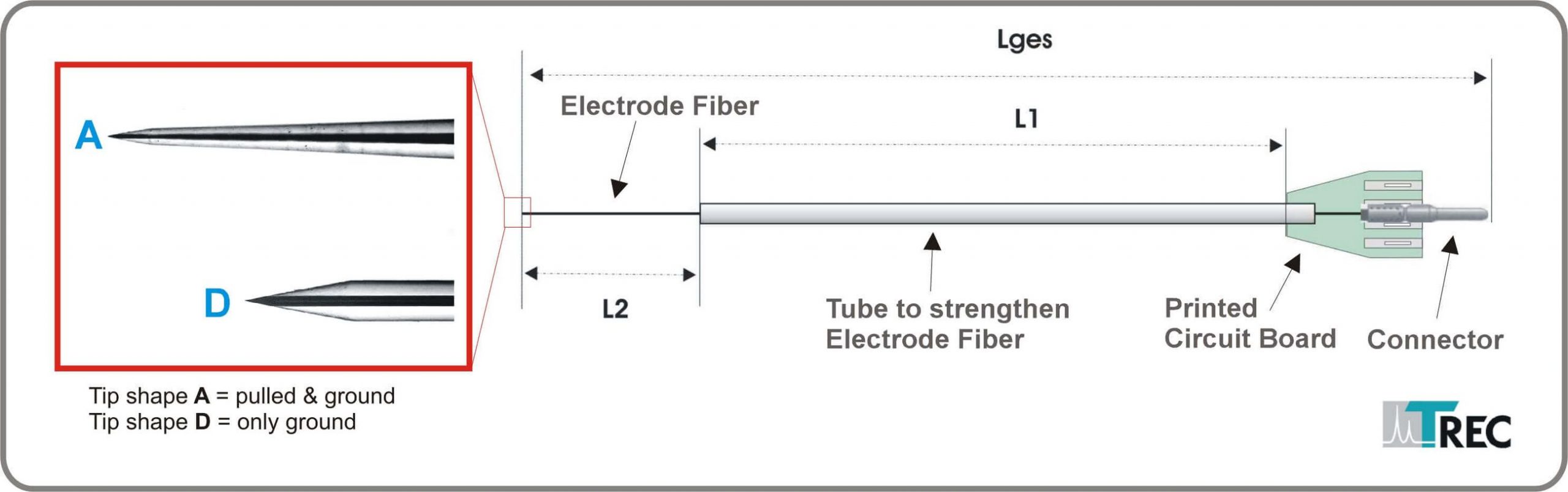
 Figure 3: Thomas microelectrodes for use with other microdrive systems
Figure 3: Thomas microelectrodes for use with other microdrive systems
Dimensions: To be specified by customer. Please see Figure 3 and/or attached Data Sheet for Thomas Electrode.
Attention: The aforementioned dimensions are determined for each Thomas microdrive system. Please provide serial number of microdrive and microdrive exchangeable head for correct electrode dimensions. If you are not sure, please contact Thomas RECORDING.
Article numbers:
AN000050 (Eckhorn Matrix, 1 piece)
AN000222 (Eckhorn Matrix, box of 12 pieces)
AN000203 (Mini Matrix, 1 piece)
AN000204 (Mini Matrix, box of 12 pieces)
AN000217 (Tetrode Mini Matrix, 1 piece)
AN000218 (Tetrode Mini Matrix, box of 12 pieces)
AN000719 (Pencil Drive, box of 12 pieces)
Overview
The electrode fibers are available with shank diameters ranging from 20 µm to 120 µm. Standard measures of our quartz-platinum/tungsten electrode fibers are 80 µm for the outer shank diameter and about 25 µm for the diameter of the metal core (other diameters on request!).
A special developed manufacturing technique makes it possible, for tips to be drawn on these thin fibers, with both the outer quartz mantle and the metal core tapering down to very small dimensions. With a specific grinding technique the metal core of every microelectrode is subsequently exposed to the required tip size respectively tip impedance (see “Manufacturing Equipment”).
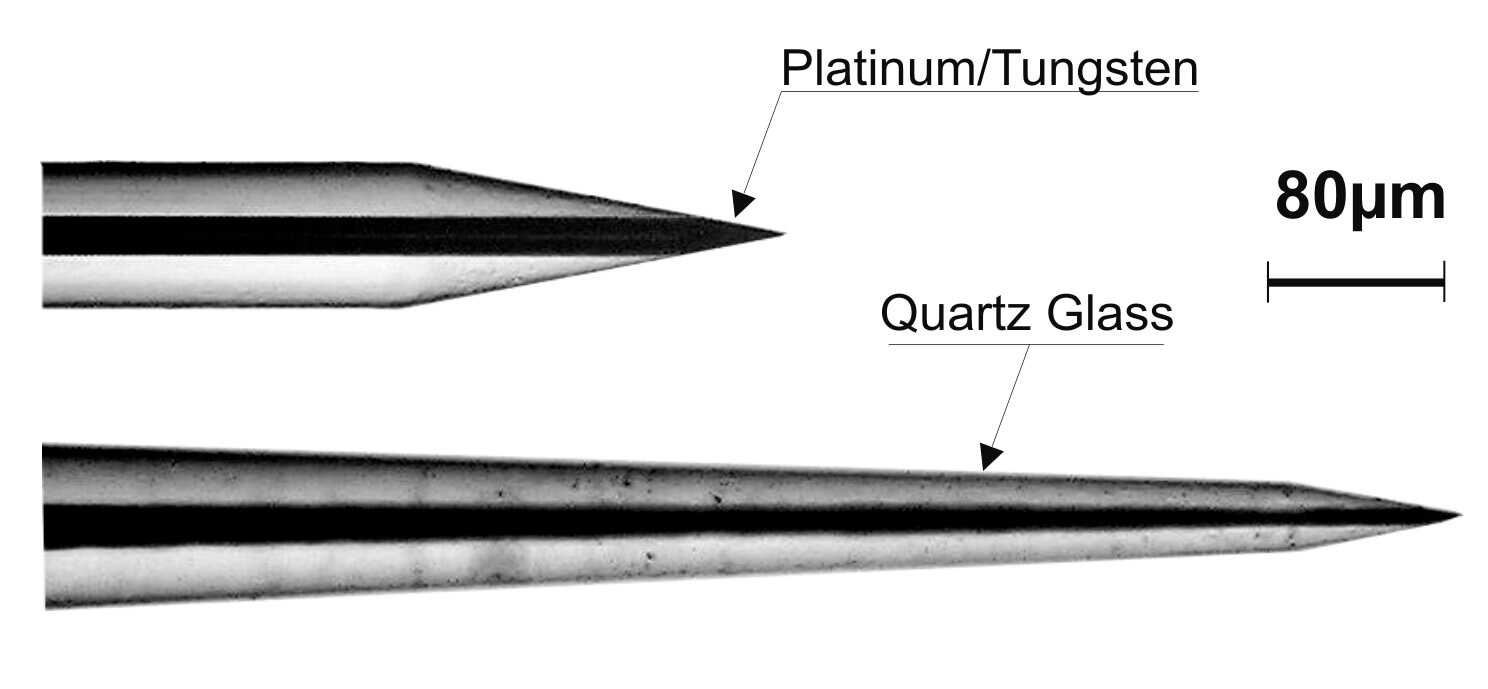 Figure 1: Different tip shapes of TREC quartz glass insulated platinum tungsten microelectrodes. Tip shape D has impedance values between 500 and 800 kΩ and is well suited for multi-unit recording. Tip shape A has impedance values between 1 and 10 MΩ (in steps of 1MΩ available) and is well suited for single unit isolation.
Figure 1: Different tip shapes of TREC quartz glass insulated platinum tungsten microelectrodes. Tip shape D has impedance values between 500 and 800 kΩ and is well suited for multi-unit recording. Tip shape A has impedance values between 1 and 10 MΩ (in steps of 1MΩ available) and is well suited for single unit isolation.
Our microelectrodes are available with two different tip profiles (see figure 1).
Electrodes with grinded tips (tip shape D) are very well suited for recording of multi-unit-activity (MUA), local-field-potentials (LFP) and for microstimulation of brain tissue (Impedance value at 1kHz in the range of 300kW to 800kW, depending on the fiber shank diameter and the grinding angle).
Electrodes with pulled and grinded tips (tip shape A), are very well suited for single-unit-recordings (Impedance value at 1kHz in the range of 1 MΩ to more than 10 MΩ, depending on the fiber shank diameter and the grinding angle).
The Quartz/platinum-tungsten fiber electrodes manufactured by Thomas RECORDING permit the recording of extracellular potentials with excellent single-unit isolation, signal to noise-ratio and temporal stability (see figure 2).
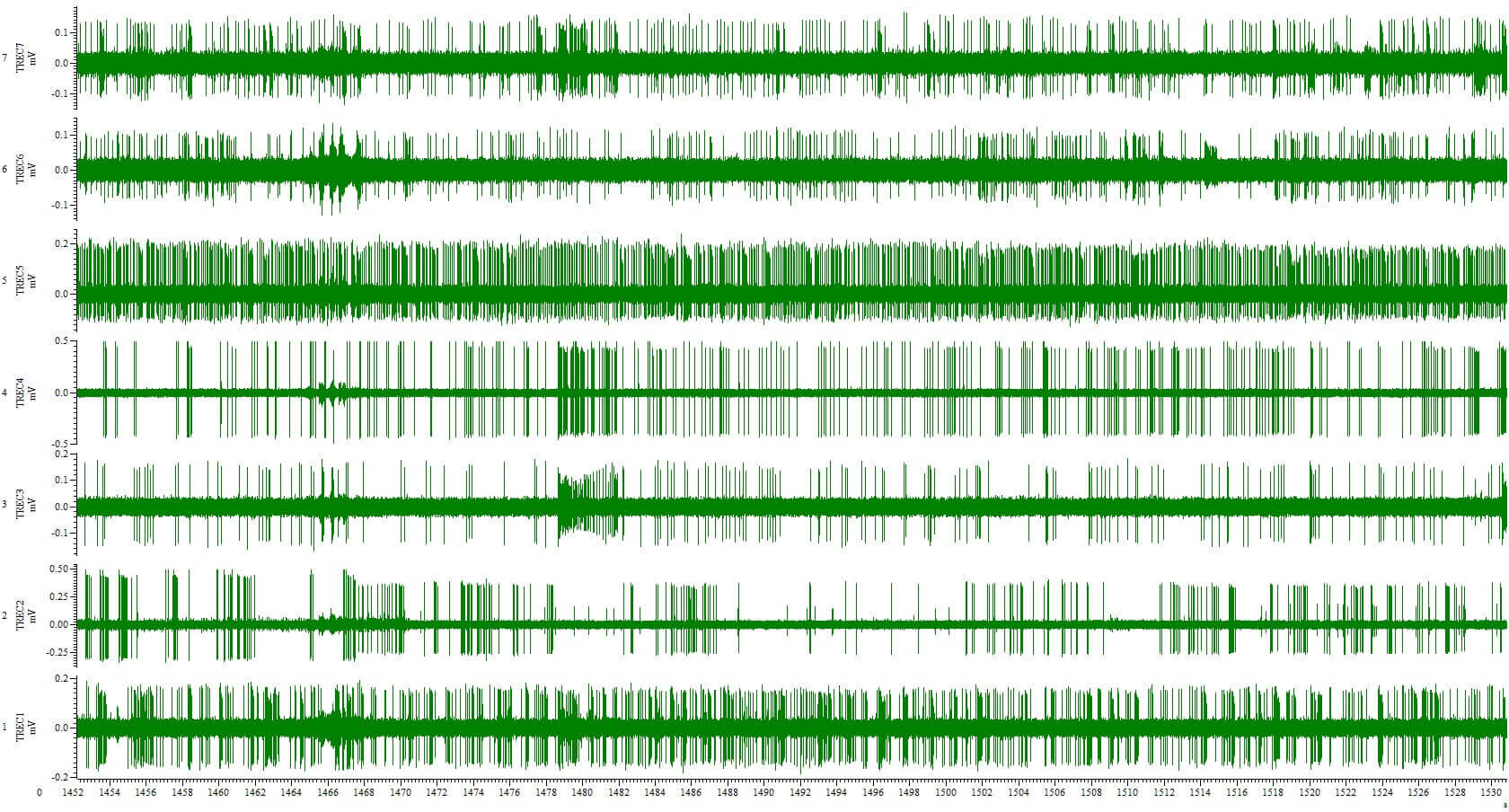 Figure 2: Extracellular recording in primate amygdala with 7 Thomas microelectrodes loaded in a TREC multielectrode drive type 7 Electrode Eckhorn Matrix. (with kind permission of Katalin M. Gothard, M.D., Ph.D.
Figure 2: Extracellular recording in primate amygdala with 7 Thomas microelectrodes loaded in a TREC multielectrode drive type 7 Electrode Eckhorn Matrix. (with kind permission of Katalin M. Gothard, M.D., Ph.D.
The method is based on a novel type of microelectrode, made of thin quartz-metal fibers. These fibers are manufactured exclusively by Thomas RECORDING GmbH using techniques adapted from optical fiber fabrication. Microelectrodes made of these fibers have outstanding recording properties:
- Quartz/platinum-tungsten fiber electrodes permit the recording of extracellular potentials with excellent single-unit isolation, signal-to-noise ratio, and temporal stability.
- The electrodes have a wide recording bandwidth and a low cut-off frequency, so that both spike potentials and (slow) local field potentials can be recorded from the same electrode.
- Due to their smooth shape and small dimensions the electrodes cause only minimal tissue damage.
- The electrodes are strong enough to penetrate the dura of a monkey or a cat. Since the dura does not need to be opened, the preparation is simplified and the danger of trauma and of infection in chronic preparations is greatly reduced.
We offer our unique quartz glass insulated platinum/tungsten microelectrodes also for use with other Microdrive systems and recording applications including custom design systems and implants.
Figure 3: Thomas microelectrodes for use with other microdrive systems
It is our strength to customize our microelectrodes to meet your requirements. Please do not hesitate to contact us for further information.
 Figure 4: Thomas microelectrodes with patented rubber tube drive loaded to a Thomas Microdrive type “7 electrode Eckhorn Matrix”
Figure 4: Thomas microelectrodes with patented rubber tube drive loaded to a Thomas Microdrive type “7 electrode Eckhorn Matrix”
Advantages of Thomas microelectrodes
Some advantages of our worldwide used microelectrodes are:
- The passage between the glass isolation and the core of all of our microelectrodes is very smooth (see figure 1 below). The result is, that there will be less tissue damage during the electrode penetration. Other electrodes like silicon-substrate electrodes or isolated metal-wire electrodes usually have cutting edges or a terraced passage between the core isolation and the tip and they will cause more tissue damage. Due to their smooth shape and their small dimensions, our microelectrodes cause only minimal tissue damage. Tissue damage by our microelectrodes, in fact, is so small that electrode tracks cannot be verified with standard histological techniques!

Figure 5: The passage between the glass insulation and the metal part of the electrode tip is so smooth, that only minimal tissue damage is caused when the electrode is introduced in brain tissue.
- Due to the geometrical shape of our fiber microelectrode (see figure 5), tissue is displaced radially during penetration, with little tissue compression. There is little or no-readjustment of tissue after insertion, which might be one of the reasons why injury potentials are hardly ever seen after electrode movement, has been stopped.
- Microgrooves, caused by the grinding process, increases the effective tip area at a given tip volume (see figure 6). This results in a tip capacitance of more than 2pF/µm2, which is considerably higher than the tip capacitance of etched tips. This high tip capacitance of our microelectrodes is one reason for their excellent signal-to-noise ratio and single-unit isolation. So our quartz-platinum/tungsten fiber electrodes permit the recording of extracellular potentials with excellent single-unit-isolation, signal-to-noise ratio and temporal stability
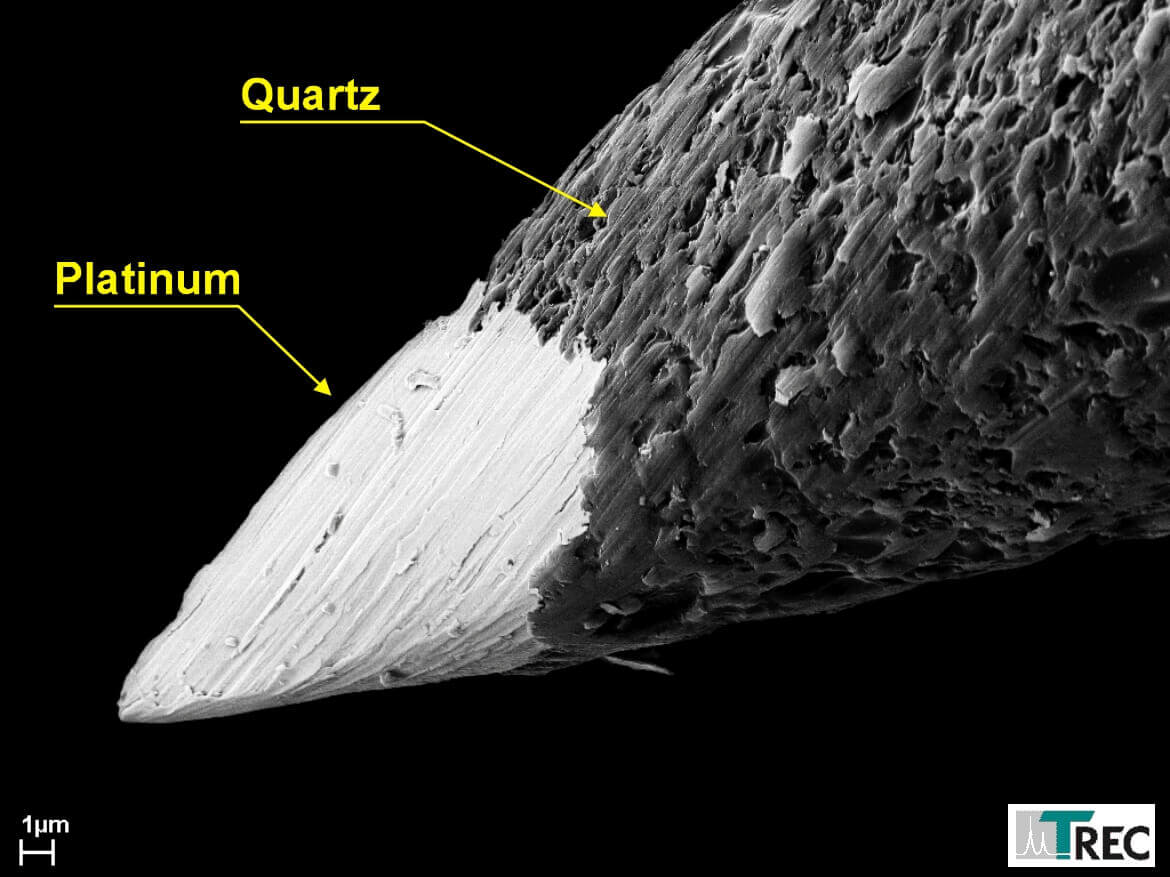 Figure 6: Microgrooves in the metal increase the effective metal area which causes a higher tip-tissue capacity, one reason for the excellent signal to noise ratio of these electrodes.
Figure 6: Microgrooves in the metal increase the effective metal area which causes a higher tip-tissue capacity, one reason for the excellent signal to noise ratio of these electrodes.
- The shaft diameter of the most conventional microelectrodes, generally, is relatively large (up to 300µm). It is therefore, not possible to use such electrodes in closely spaced parallel arrays for multi-channel recordings. The shaft of our microelectrodes, over the whole penetration depth, is cylindrical and very thin (see figure 7). Our standard microelectrode has a shaft diameter of 80µm. Smaller diameters down to 20µm are also available. So our fiber microelectrodes are well suited for multi-electrode arrays. In contrast to silicon-substrate electrodes (Wise and Angell, 1969; Wise and Starr, 1969) movement control of individual electrodes is possible.
 Figure 7: Shaft diameter of Thomas fiber electrodes is small and constant over the complete length of the electrode fiber.
Figure 7: Shaft diameter of Thomas fiber electrodes is small and constant over the complete length of the electrode fiber.
- Our microelectrodes have a wide recording bandwidth and a low cut-off-frequency, so that both, spike potentials and (slow) local filed potentials can be recorded from the same microelectrode.
- The electrodes are strong enough to penetrate the intact dura of monkey or cat for a relatively long time. Since the dura does not to be opened, the preparation is simplified and danger of trauma and of infection in chronic preparations is greatly reduced.
- If our electrodes are used together with Thomas Microdrive systems, it is possible to drive the electrodes over long distances through the brain (e.g. 40mm). The electrodes will move straight and do not leave the trajectory to the target (e.g. primate amygdala). See figure 8.
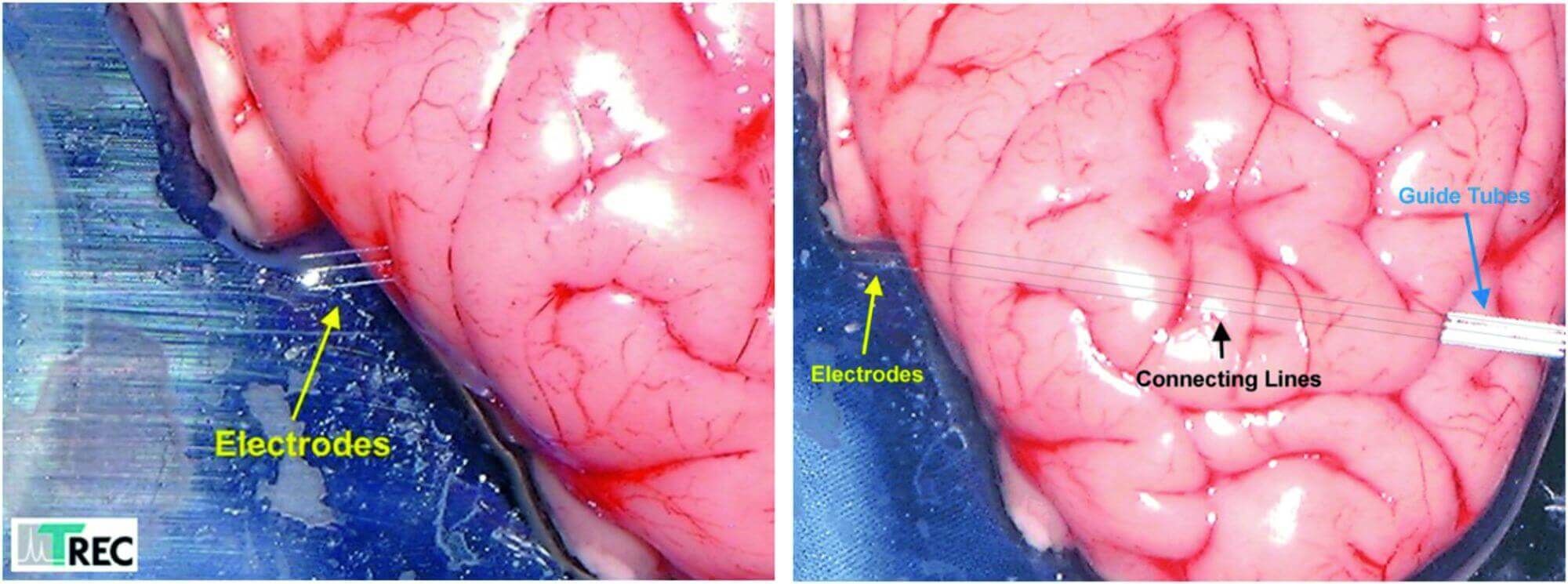 Figure 8: These pictures show a penetration test with Thomas fiber microelectrodes. The electrodes moves straight through the brain over a distance of 40.000µm although the fibers just had 80µm outer diameter. The drawn connecting lines in the rigth photo show that the exit points of the elelctrodes are exactly at the opposite side of the entrance points.
Figure 8: These pictures show a penetration test with Thomas fiber microelectrodes. The electrodes moves straight through the brain over a distance of 40.000µm although the fibers just had 80µm outer diameter. The drawn connecting lines in the rigth photo show that the exit points of the elelctrodes are exactly at the opposite side of the entrance points.
- Each of the four platinum-tungsten-cores of our multifiber electrodes are totally isolated from the other cores because of the good isolation property of quartz glass. This avoids any cross-talk between the cores.
- Because of our precision manufacturing techniques the geometrical shape of the electrode tip can be made exactly and reproducibility according to specifications, as long as it is a tip geometry that is grindable. Conical tips can be reproduced with high accuracy (see figure 7). With our precision manufacturing equipment tip geometry of the microelectrodes can be optimized for a particular recording situation. Each microelectrode will be double checked, electronically and microscopically. We give the impedance value for each electrode, documented in a test certificate, enclosed to the electrode delivery.
- We are able to produce microelectrodes that will meet the special requirements of our customers. For example different electrode lengths, different tip profiles, special impedance values (for single-unit isolation, multi-unit isolation, local field potential recordings or stimulation) are available. Please ask for your special configuration.
- Electrode tip manufacturing equipment is also available from Thomas RECORDING GmbH. With this equipment the scientist will be able to manufacture his own microelectrodes for his special neurophysiological application.
- Our preamplifiers and filter systems are especially developed for use with our microelectrodes. So complete recording systems with optimal adaptation to a special neurophysiological application are available.

NEWS

PRODUCTS

SOLUTIONS

DISTRIBUTORS
 Sign Up
Sign Up